
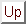

The key long-range model results depend on the properties of the local cortical circuitry, so we begin by reviewing and expanding on these results. We have recently described a local circuit model of orientation selectivity in V1, which accounts for the results of most intracellular, extracellular and pharmacological studies of orientation selectivity [38]. This model requires only weakly oriented thalamic inputs and utilizes recurrent, short-range excitatory connections between neurons to amplify and sharpen orientation selectivity. Inhibition contributes indirectly to tuning by balancing excitation within the columnar population. In this model cortical neurons retain sharp orientation tuning independent of stimulus contrast [1,14,35,36]. This physiological property cannot be achieved solely by the broadly oriented thalamocortical inputs used in the model (see [38]), but rather requires cortical inputs. Typical contrast response curves generated by the model (figure 1) closely resemble experimental curves from V1 neurons [1,4,11]. One key property of this cortical module is that cortical excitation amplifies low contrast responses, while cortical inhibition grows strong at high contrasts to attenuate responses (e.g., contrast saturation).
Thus this cortical circuitry not only provides sharp orientation tuning but also yields a dynamic gain control mechanism: recurrent excitation strongly amplifies preferred responses at low levels of thalamic drive while inhibition reduces amplification or even attenuates preferred responses at high levels of thalamic drive. Several differential properties of excitatory and inhibitory neurons and synapses contribute within a local feedback circuit to generate this gain regulation as a network property. Fast-spiking inhibitory interneurons exhibit higher firing rates and greater frequency vs. current slopes, due to the higher input resistance and brief after-hyperpolarization potentials of inhibitory neurons [27]. Inhibitory neurons due to their lower surface areas appear to receive fewer synaptic inputs and thus may have higher contrast thresholds. Other possible differences between excitatory and inhibitory neurons may also exist, but have not been considered here. Such a variable gain control mechanism appears well suited to contribute to other reported cortical response nonlinearities within the classical receptive field. For example, cortical neuron responses decline over a broad range of stimulus velocities for which LGN responses increase [30].

Figure 1: Cortical gain control of local circuitry.
Response amplitude
is shown as a function of stimulus contrast for an uncoupled cortical
cell (dashed red line) and for a cortical cell with short-range cortical
excitatory and inhibitory connections (solid blue line). Cortical connections
generate a contrast-response function with greater slope (high gain) at
low contrasts and stronger response saturation (low gain) at high contrasts
than the uncoupled cell.
Cortical gain control arising from local connections is also the substrate for receptive field dynamics that arise from long-range horizontal connections. We have constructed a 1:40 scale model of the cortical circuitry under a 2.5 mm by 5 mm region of cortical surface. The model contains over 20,000 spiking neurons with approximately 1 million synaptic connections. (A preliminary version has been reported previously [37]).

Figure 2:
Long-range and short-range connectivity of the model.
Columnar orientation preferences are shown on the
color map (see key). Mesh elevation represents the strength
of total synaptic connections ( Excit - Inhib ) to cells of the central
(green) orientation
column. Long-range synapses (excitatory only) generally connect
domains with similar orientation preference. Short-range connections
consist of both excitatory
and inhibitory inputs with excitatory inputs stronger at shortest distances
and inhibitory inputs stronger at greater distances.
Long-range connections originate from excitatory neurons and target both excitatory and inhibitory neurons with similar orientation preferences (see methods; figure 2). Large scale computer simulations were employed to compare neuronal responses to oriented grating stimuli that covered (a) the classical receptive field alone (``center'' stimuli), (b) the classical and extraclassical receptive fields (``center + surround'' stimuli). Center contrast in both cases was varied systematically from zero to high, while the iso-orientation surround grating was kept at high contrast. The surround grating modulated center responses in a way that depended on the contrast of the center grating. Responses to low contrast center gratings were facilitated by the presence of a high contrast surround grating, while responses to high contrast center gratings were suppressed by the same high contrast surround grating (figure 3). The surround stimulus, and thus the long-range inputs, were the same for both low and high contrast centers; however, these inputs had opposite effects due to the gain control state of the local cortical circuitry. Thus, the model predicts that fixed-strength long-range connections can have bi-phasic modulatory effects, depending on the strength of the central excitatory drive. Although both kinds of modulatory effect could be produced by surround gratings that differed in orientation from the center grating, due to weaker long-range inputs the effects were substantially weaker than for iso-orientation gratings (simulation results not shown).

Figure 3: Biphasic surround modulation.
Average response amplitude of 144 excitatory neurons
in the central iso-orientation domain is shown as a function of center
stimulus contrast. Solid blue line is response to center stimulus alone;
dashed red line is response to center plus high contrast surround stimulus.
The presence of the iso-orientation surround stimulus facilitates responses
to low contrast center stimuli, but suppresses responses to high contrast
center stimuli.