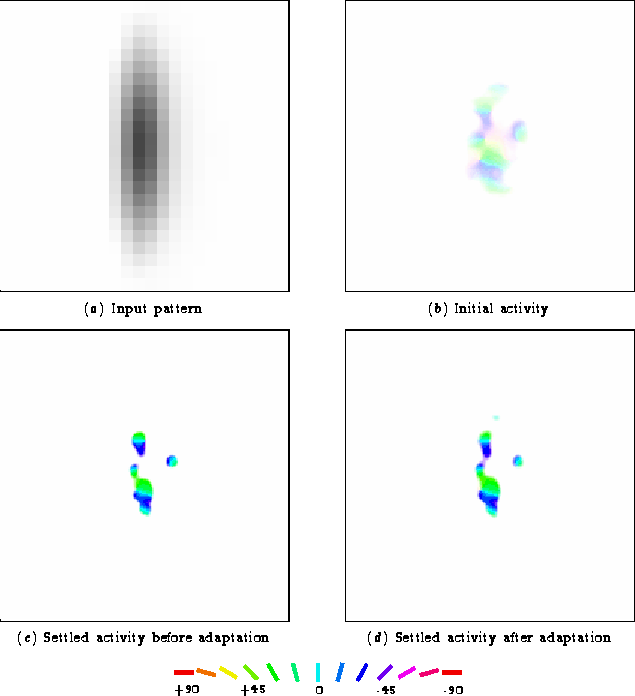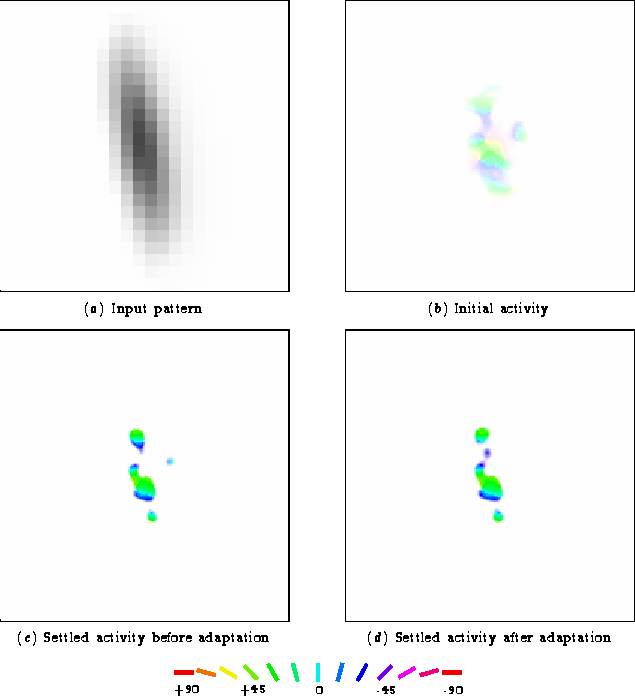Figure 5.6 shows the response to the adaptation
line before and after adaptation.
The sharpening is caused by the strengthening of inhibition between activated neurons -- only the areas with the strongest activation remain active after settling. The rest of the neurons initially activated in figure 5.6b are silenced by the active ones in the center. The sharpening effect is present even before adaptation, which is why the settled activity differs from the initial activity, but it gets stronger with adaptation. The inhibitory weight changes and the activity changes can be related to each other by comparing the areas showing changes between figure 5.6c and 5.6d with the areas showing increased inhibition in figure 5.5a. The increased inhibition has effectively created a group of competitive neurons where only a central region of strongly activated neurons is allowed to continue responding.
None of these changes show up in the TAE curve in figure 5.3 at 0° because the distribution of activated units contains approximately the same proportion of each orientation preference both before and after adaptation. Compared to figure 5.6c, there are more green units (orientation preference less than 0°) activated in the center of figure 5.6d. At the same time, there are more blue units (orientation preference greater than 0°) activated just outside that area. These changes cancel out for the adaptation orientation, since they are centered upon it. As discussed in the next section, the direct TAE occurs for test lines of slightly different orientations, where the changes do not cancel out.
For the peak of the direct effect, the perceived orientation of a
10° test line shifted away from 0° by several degrees
(figure 5.3). This change is quite
subtle but evident in the activity patterns for the +10° test
line in figure 5.7.
The changes are once again due to the strengthening of the inhibitory connections. As shown in figure 5.5a, the inhibitory connections between the green, cyan, and blue neurons were strengthened. This test line initially activated the green orientations most strongly, so lateral interactions allowed those neurons to silence the weakly-activated cyan and blue areas. The increased inhibition between neurons active during adaptation results in competition between them for activation -- only a subset will remain active at a given time. Because these changes occurred only for neurons preferring orientations closer to 0° than 10°, the perceived orientation shifts away from 0°. Thus direct tilt aftereffects are occurring in the RF-LISSOM model exactly as predicted by the lateral inhibition theory.
 |
The indirect effect, true to its name, results only indirectly from the strengthening of the inhibitory connections. The inhibition between the green, cyan, and blue neurons was strengthened during adaptation. Of those, only the green neurons were activated by this test line, since it is quite distant from the adaptation line. Thus the stronger inhibitory connections themselves did not change the response pattern significantly, since the targets were already inactive.
However, since the total inhibitory connection strength to each neuron is limited (equation 3.4), the increases in strength among the green, cyan, and blue neurons come at the expense of all the other inhibitory connections to the green neurons. As shown in figure 5.5b, the weakened connections are spread out over neurons with a wide range of orientation preferences. The weakened connections include those to nearly all of the units activated for this test line. Thus those units inhibit the green areas much less than they did before adaptation. Consequently, more of the green neurons remain active throughout the settling process. Before adaptation, many of them had been silenced by the other neurons closer to the orientation of the test line. The net effect of these changes is that the perceived orientation shifts towards 0°.
The magnitude of the indirect effect is smaller than the direct effect because fewer adapted neurons are involved. Most of the neurons showed little change; only those few closest to the adaptation orientation were affected. For the direct effect, many neurons were affected -- a number of those preferring orientations greater than that of the test line became less active, and some of the ones preferring smaller orientations became more active. These changes add up to a greater difference in the perceived orientation for the direct effect than for the indirect effect.
The indirect effect in RF-LISSOM is a simple consequence of the normalization of inhibitory weights after they have been adjusted by Hebbian learning. This explanation for the indirect effect was not discovered until the TAE was modeled here, thus underscoring the importance of building computational models. Chapter 6 will discuss the biological evidence for this theory of the indirect effect, and will compare it to the evidence for other theories that have been proposed.
The model has allowed us to look into the cortex as it is processing and see the changes as they occur. No other computational model has allowed the tilt aftereffect to be studied so concretely. As also discussed in chapter 6, it may be possible to obtain similar plots for the cortex to compare with these results.